STATISTICAL ANALYSIS OF EVOLUTIONARY TRENDS
|
|||||
|
Introduction Disclosure of findings and
contributions to human resources |
Evolutionary Trends in Growth RatesWe use several empirical methods that optimize calculated growth rates
and histological patterns on existing phylogenetic hypothesis. These
methods allow us to test hypotheses regarding where advance growth rates
first appear in the coelurosaur family tree and whether avialan growth
rate increase occurred via a single event, multiple events, or spanned
several phylogenetic nodes. Three main approaches exist among all available
methods to optimize quantitative data, such as growth rates, on a phylogenetic
tree: linear parsimony (Farris, 1970; Swofford and Maddison, 1987),
squared-change parsimony (Huey and Bennet 1987, Maddison, 1991, McArdle
and Rodrigo 1994), and model based methods (Martins and Hansen 1997,
Schluter et al. 1997). These methods differ both on the assumptions
and on the information that can be retrieved from them. We employ linear
parsimony and squared-change parsimony since because they have been
thoroughly tested. |
||||
|
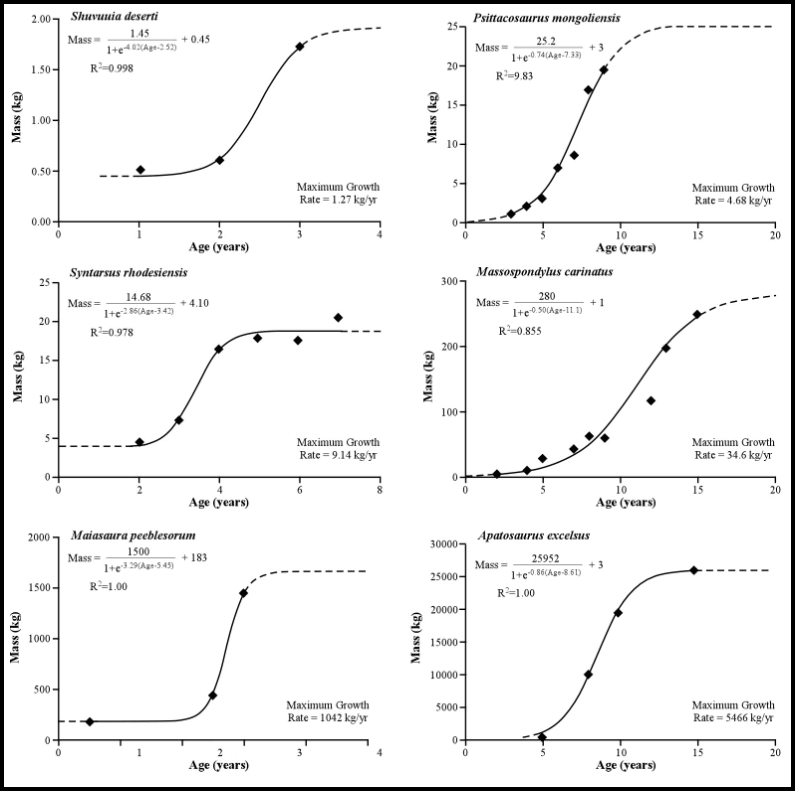
Findings to Date Our studies are demonstrating that histological assessments of longevity coupled with DME derived body mass quantification can be used to accurately reconstruct growth curves from the fossilized remains of dinosaurs (Erickson and Tumanova 2000, Erickson et al. 2001; Fig. 6). From these curves the temporal length of the lag, exponential, and stationary stages of development can be elucidated, and exponential stage growth rates and ages at attainment of somatic maturity can be quantified.
|
|||||
| Figure 6. Growth curves for non-avian dinosaurs assessed from femoral histology and DME (From Erickson, Curry Rogers, and Yerby 2001). Our analysis revealed that sigmoidal equations accurately described the growth data for the six dinosaurs we tested (r 2 = 0.885 - 1.0). Exponential stage growth rates ranged from 3.4 to 14,460 g/day with values positively correlating with increased adult mass. No taxon substantially deviated from the general dinosaurian trend (r2 = 0.96). The length of the exponential growth stages ranged from approximately one to six years. The onset of somatic maturity occurred between the ages of three and thirteen with values positively correlating with increased body size. | |||||
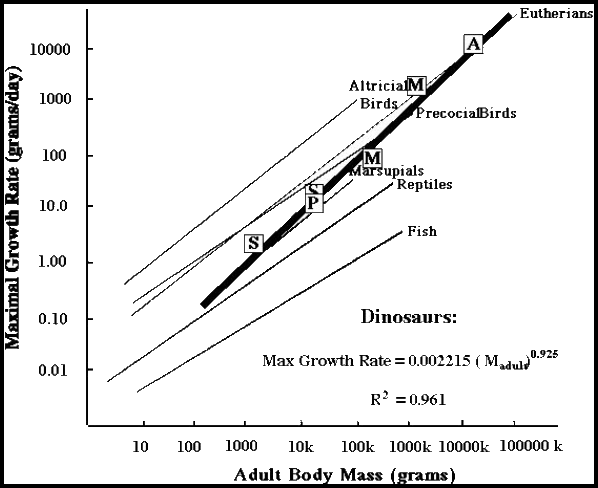
| Our sample of phylogenetically disparate dinosaurs have shown that all exhibited exponential stage growth rates higher than those found in traditional "reptile" groups. In addition, a unique general "dinosaurian" growth pattern has been discovered (Erickson et al. 2001, Fig. 7). Our testing of close outgroups to birds is showing rates typical of non-avian dinosaurs and no-where near as high as those in living birds. Hence we have evidence that nearly the entire evolution to rapid precocial rates occurred in taxa closer to the Avialae or after the evolution of birds. | |||||
|
|
|||||
| Figure 7. Exponential growth of non-avian dinosaurs compared with data for living vertebrate groups (from Erickson et al., 2001). The smallest dinosaurs (<1.0 kg) are predicted to have growth rates 5-8 times less than the more rapid rates for precocial avialans. DME assessments for dinosaurs from new histological data of Erickson et al. (2001); Chinsamy (1990, 1993); Erickson and Tumanova (2000); Curry (1999); and Horner et al. (2000). Data for extant taxa from compilations by Case (1978), Calder (1984). Progressing smallest to largest: S = Shuvuuia, P = Psittacosaurus, S = Syntarsus, M = Massospondylus, M = Maiasaura, A = Apatosaurus. The regression line extends to the range of size for non-avian dinosaurs (0.2 kg - 100,000 kg). | |||||
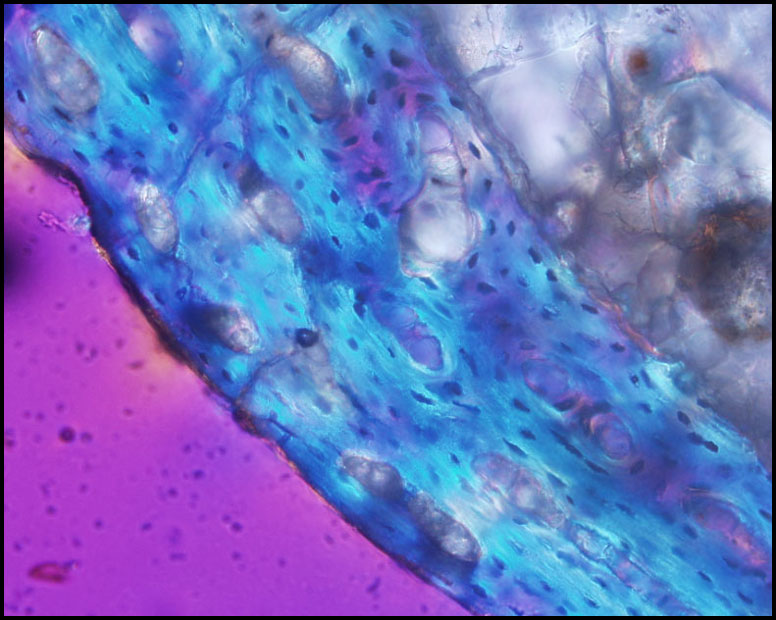
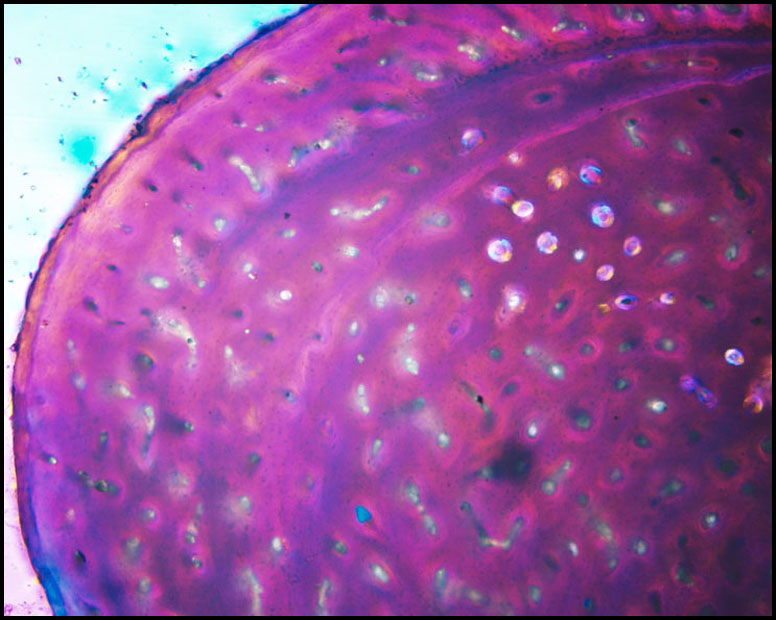
Recently processed specimens: |
|||||
|
Byronosaurus femur-Hatchling
|
|||||
|
Conchoraptor fibula
|
|||||
|
Conchoraptor hatchling fibula
|
|||||
|
Deinonychus gastralia
|
|||||
|
Citpati femur
|
|||||
|
Shuvuuia femur
|
|||||
|
Tyrannosaurus rex rib
|
|||||
|
Gorgosaurus rib
|
|||||
|
Velociraptor fibula
Next: Roles of the P.I.'s |
|||||