| Phylogenetic relationships and species limits within Phyllotis (Rodentia: Sigmodontinae): concordance between mtDNA sequence and morphology. Steppan, S.J. 1998. Journal of Mammalogy |
| Phylogenetic relationships and species limits within Phyllotis (Rodentia: Sigmodontinae): concordance between mtDNA sequence and morphology. Steppan, S.J. 1998. Journal of Mammalogy |
Abstract
Phylogenetic relationships and species limits within the South American rodent genus Phyllotis are estimated using nucleotide sequence data from a 973 base-pair region of the mitochondrial gene cytochrome b. Geographic variation and species limits are assessed by sequencing individuals from multiple populations and by morphometric analyses of museum skulls and skins. The results support a clade including Phyllotis and Auliscomys but not Graomys, the darwini species group, and a northerly distributed species-pair of P. amicus and P. andium. The phylogeny, genetic divergence estimates, and morphology indicate that P. xanthopygus rupestris is polyphyletic, and that populations from the Pacific slope of the Andes currently assigned to that taxon are specifically distinct from P. xanthopygus. These populations are placed in the newly elevated P. limatus and an emended diagnosis is provided. The biogeographic and phylogenetic patterns are most consistent with successive colonization of the Pacific slopes, possibly by range expansion and contraction of P. xanthopygus over a short period of time relative to the age of the clade.
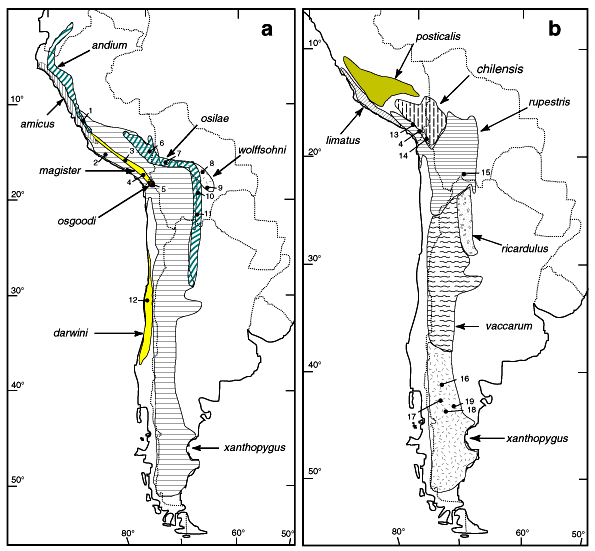
Figure 1. a) Geographic distribution of Phyllotis species sequenced for this study. b) Geographic distribution of Phyllotis xanthopygus subspecies. Numbers indicate the localities: 1, Lima, Peru; 2, Atiquipa, Caraveli, Arequipa, Peru; 3, Chivay, Arequipa, Peru; 4, Tarata, Tacna, Peru; 5, Parinacota, Tarapacá, Chile; 6, Ilave, Puno, Peru; 7, Miña Lourdes, La Paz, Bolivia; 8, Quiñe, Santa Cruz, Bolivia; 9, Padilla, Chuquisaca, Bolivia; 10, Tarabuco, Chuquisaca, Bolivia; 11, Serrania Sama, Tarija, Bolivia; 12, Fray Jorge, Coquimbo, Chile; 13, Torata, Moquegua, Peru; 14, Chapiquina, Tarapacá, Chile; 15, Iscayachi, Tarija, Bolivia; 16, Comallo, Rio Negro, Argentina; 17, Cerro del Viento, Chubut, Argentina; 18, Pampa de Agnia, Chubut, Argentina; 19, Cerro Rosario, Chubut, Argentina.
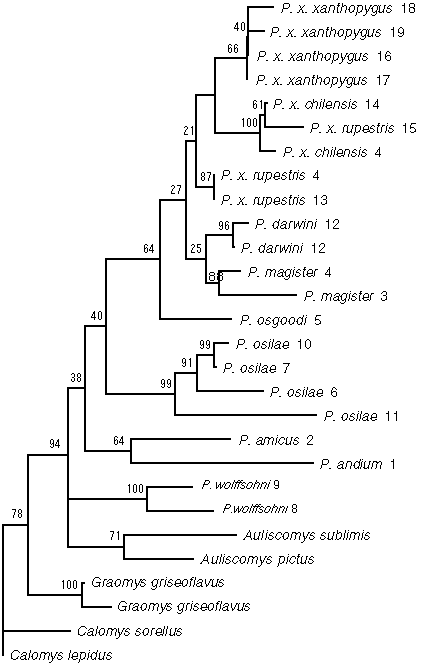
Figure 2. Strict consensus of the six most-parsimonious trees from the preferred 6-parameter analysis. Each tree is 3137 steps long. Numbers along the branches indicate the percentage of bootstrap replicates including the designated nodes. Branch lengths proportional to mean number of character state changes. Numbers following species names indicate the localities: 1, Lima, Peru; 2, Atiquipa, Caraveli, Arequipa, Peru; 3, Chivay, Arequipa, Peru; 4, Tarata, Tacna, Peru; 5, Parinacota, Tarapacá, Chile; 6, Ilave, Puno, Peru; 7, Miña Lourdes, La Paz, Bolivia; 8, Quiñe, Santa Cruz, Bolivia; 9, Padilla, Chuquisaca, Bolivia; 10, Tarabuco, Chuquisaca, Bolivia; 11, Serrania Sama, Tarija, Bolivia; 12, Fray Jorge, Coquimbo, Chile; 13, Torata, Moquegua, Peru; 14, Chapiquina, Tarapacá, Chile; 15, Iscayachi, Tarija, Bolivia; 16, Comallo, Rio Negro, Argentina; 17, Cerro del Viento, Chubut, Argentina; 18, Pampa de Agnia, Chubut, Argentina; 19, Cerro Rosario, Chubut, Argentina.
![]()
Figure 4. Elevational transects for incisor shape (depth/width) in two valleys in southern Peru: above Tarata, Tacna and above Torata, Moquegua. The Peruvian populations of rupestris are removed to P. limatus.
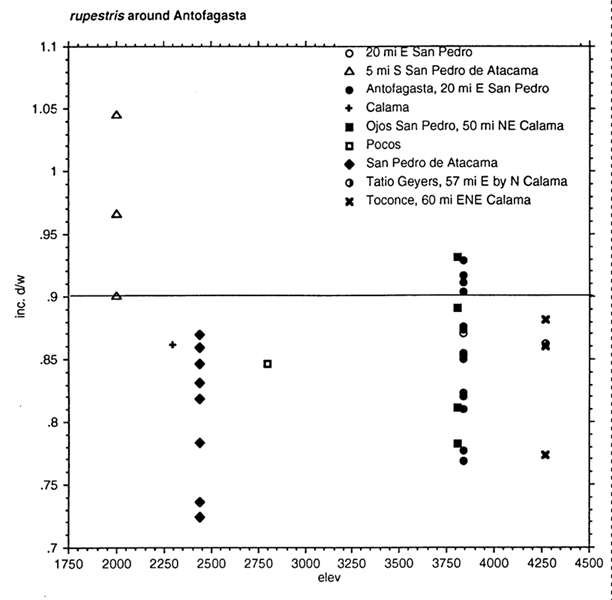
Unpublished figure: Graph of incisor shape by elevation for individuals of "rupestris" or limatus in Antofagasta, Chile.
SYSTEMATICS
Phyllotis limatus Thomas
Phyllotis darwini limatus Thomas, 1912:407, Ann. Mag. Nat. Hist.
Phyllotis xanthopygus rupestris (Gervais), 1841:115, Bull. Soc. Philom., Paris. (part)
Holotype. -- Skin and skull of an old male, British Museum (BMNH) no. 0.5.7.43, collected 29 January, 1900 by P. O. Simons, at Chosica, Dept. Lima, Peru, 3000 ft.
Emended diagnosis. -- A member of the darwini species group that can be distinguished from other members by deep and narrow upper incisors (depth at midarc greater than 90% of width across both incisors at the cutting edge), short to moderate molar row (4.2-5.8 mm), light coloration, belly frequently white, two pairs of preputial glands, and karyotype 2n=38.
Geographic distribution. -- Arid coast and Pacific slopes of the Andes in southern Peru and northern Chile from sea level to 2500 m in the north and seal level to 4000 m in the south (Fig. 5): departments of Arequipa, Ayacucho, Huancavelica, Ica, Lima, Moquegua, and Tacna in Peru, provinces of Antofagasta and Tarapacá (San Pedro de Atacama) in Chile.
Taxonomy. -- The distinctive nature of subspecies limatus relative to "rupestris" has been well accepted. Even Hershkovitz (1962) , who employed very inclusive species and subspecies concepts, retained limatus as a distinct subspecies. The tails of subspecies limatus have been described as relatively longer than those of "rupestris" (Hershkovitz, 1962; Pearson, 1958) , but while the are significantly longer on average (115% body length vs. 105%), the variation appears to be clinal and is not a diagnostic character at the individual level (Steppan, unpubl. data). Tails of limatus often have white tips, a rare trait in Phyllotis, but the feature is not universal. On the other hand, length of the auditory bulla (associated with ear size) shows a step-cline (Fig. 6). Using the break in the step cline to define the boundary between these sets of populations moves the coastal section of the boundary 250 kilometers to the northwest of its historical location (cf. Hershkovitz, 1962) No name for these Pacific slope "rupestris" populations currently exists, and further research may indicate that subspecific status is warranted. The distinctiveness of limatus may have been overemphasized by previous workers because they compared it to a composite taxon, "rupestris."
{kind=link}